
Ponerinae, Formicidae, Hymenoptera, Insecta, Arthropoda, Animalia
John T. Longino
The Evergreen State College
Olympia WA 98505 USA
longinoj@evergreen.edu
13 August 1997
| Key to Costa Rican species of Simopelta | Species List |
Taxonomy of the genus
Mayr (1870) established the genus Belonopelta for B. attenuata (type species) from Colombia. Belonopelta curvata Mayr 1887 (from Brazil), B. pergandei Forel 1909 (from Guatemala), and B. jeckylli Mann 1916 (from Brazil) were subsequently described. Mann (1922) described B. deletrix from Honduras, placing attenuata and deletrix in Belonopelta sensu stricto, and curvata, pergandei, and jeckylli in a new subgenus Simopelta. Wheeler (1935) raised Simopelta to generic status, and described new species. Borgmeier (1950), Gotwald and Brown (1967), and Snelling (1975) followed this scheme and described new species. Baroni-Urbani (1975) synonymized Simopelta and Emeryopone under Belonopelta, and placed B. deletrix in a new genus Leiopelta. Brandao (1989) followed Baroni-Urbani's scheme, naming a new species from Brazil. Holldobler and Wilson (1990) resurrected Simopelta and synonymized Leiopelta under Belonopelta. I examined the types of B. attenuata in the Mayr collection and concur that it is clearly related to B. deletrix, and nothing like Simopelta.
Natural History
Simopelta is common in mid-montane wet forest (500-2000m), and rare or absent in lowland forests. Workers are tolerant of cold wet conditions, and may be observed foraging in cloud forest during inclement weather. They forage on the ground and in the canopy.
Simopelta has behavior and morphology convergent with Ecitoninae (Gotwald & Brown 1967). Workers forage in dense columns. Raiding columns repeatedly have been observed attacking Pheidole nests. When colonies are found with brood (their own, not prey), the brood is generally of uniform size, suggesting synchronized brood development. Colony emigrations are encountered with workers transporting brood and the queen traveling with the column, suggesting nomadic behavior. Workers have small eyes, reduced to a single facet. The queen is "dicthadiiform" (permanently wingless, with grossly elaborated petiole and swollen gaster) like Ecitoninae.
Nest sites are variable and reveal additional similarities to ecitonine behavior. Temporary nest sites during colony emigrations have been observed twice, once in a hollow dead stick suspended in low vegetation (Gotwald & Brown 1967), and once under a rock. Apparent longer-term nest sites have been observed twice, in extensive chambers in soil. In one of these observations brood were dispersed in various chambers, along with prey remains. It is unknown whether the Simopelta excavated the chambers themselves or occupied preexisting chambers, but the behavior suggests periods of stable use of one nest site, similar to the statary phase seen in Ecitoninae. Males of Simopelta are unknown.
If males were also convergent on Ecitoninae, one would expect them to be conspicuous and robust, sausage-shaped, and with strong mandibles, and they would appear at lights and/or in malaise traps. Simopelta may have no males. If this is true, and the colonies reproduce by budding (similar to Ecitoninae), then the different "species" of Simopelta could be distinct clonal lineages.
The genus is known from scattered montane localities throughout the neotropics (southeastern Brazil, Peru, Ecuador, Costa Rica, Guatemala). Given the high diversity within Costa Rica, and the possibility of clonal reproduction, we might expect high regional differentiation wherever the genus is found.
Several features of their biology result in their being infrequently collected. They are most abundant in a spatially restricted and less frequently visited habitat: mid-montane wet forest. Like Ecitoninae, they have large colonies and are low density relative to other ants. They are best encountered not by intensive searching in a small area (typical ant collecting behavior) but by walking trails looking for raiding columns.
Their restricted habitat preference and low density should make them particularly prone to local extirpation from a region, and this may have happened already for at least one species in Costa Rica (pergandei). If Simopelta shows great regional differentiation, and the species have narrow geographic ranges, it is likely that widespread extinction of species has already occurred and will continue.
Literature Cited (this page and species accounts)
Baroni-Urbani, C. 1975. Contributo alla conoscenza dei generi Belonopelta Mayr e Leiopelta gen. n. (Hymenoptera: Formicidae). Mitt. Schweiz. Entomol. Ges. 48:295-310.
Borgmeier, T. 1950. A femea dichthadiiforme e os estadios evolutivos de Simopelta pergandei (Forel), e a descricao de S. bicolor, n. sp. (Hym. Formicidae). Rev. de Entomologia 21:369-380.
Brandao, C. R. F. 1989. Belonopelta minima new species (Hymenoptera, Formicidae, Ponerinae) from eastern Brazil. Rev. Bras. Entomol. 33:135-138.
Forel, A. 1909. Ameisen aus Guatemala usw., Paraguay und Argentinien (Hym.). Dt. Entomol. Z. 1909:239-269.
Gotwald, W. H., and W. L. Brown Jr. 1967 (1966). The ant genus Simopelta (Hymenoptera: Formicidae). Psyche 73:261-276.
Holldobler, B., and E. O. Wilson. 1990. The Ants. Harvard University Press, Cambridge, Massachusetts, USA.
Mann, W. M. 1916. The Stanford Expedition to Brazil, 1911, John C. Branner, Director. The ants of Brazil. Bull. Mus. Comp. Zool. 60:399-490.
Mann, W. M. 1922a. Ants from Honduras and Guatemala. Proc. U.S. Natn. Mus. 61:1-54.
Mayr, G. 1870. Formicidae novogranadenses. Sitz. Akad. Wiss. Mathem.-naturwiss. 61:370-417.
Mayr, G. 1887. Sudamerikanische Formiciden. Verh. Zool.-bot. Ges. 37:511-632.
Snelling, R. R. 1971. A new species of Simopelta from Costa Rica (Hymenoptera: Formicidae). Bulletin S. Calif. Academy of Sciences 70:16-17.
Wheeler, G. C., and J. Wheeler. 1986. Supplementary studies of ant larvae: Ponerinae. Trans. Amer. Ent. Soc. 112:85-94.
Wheeler, G. C., and J. Wheeler. 1989. Notes on ant larvae: Ponerinae. J. N. Y. Entomol. Soc. 97:50-55.
Wheeler, W. M. 1935. Ants of the genera Belonopelta Mayr and Simopelta Mann. Rev. de Entomologia 5:8-19.
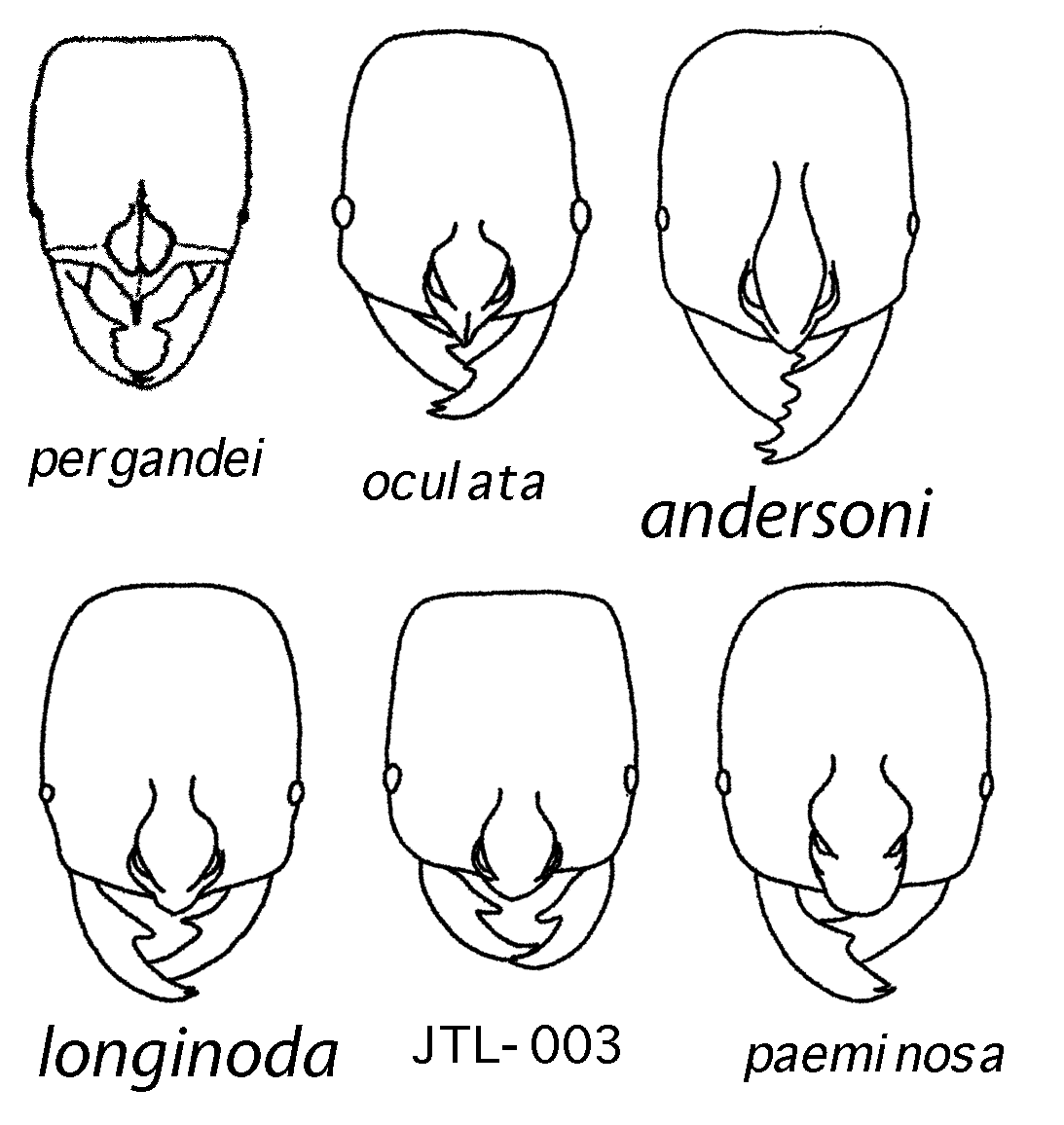
Illustrations of characters and discussion of character variation among the Costa Rican species follow the key.
1a. Mandibles with teeth or denticles between the large basal tooth and the two large apical teeth; no katepisternal tooth: 2
1b. Mandibles with no teeth or denticles between the large basal tooth and the two large apical teeth; katepisternal tooth present: 3
2a. Anteromedian clypeal border with spine: pergandei
2b. Anteromedian clypeal border triangular: JTL-001
3a. Eye relatively large; mandible relatively narrow with acute basal tooth; face mainly punctate, in lateral view with an abrupt transition to sublinear, transverse rugae; anteromedian clypeal margin subtriangular: oculata
3b. Eye relatively small; mandible narrow or broad, with basal tooth acute or blunt; face punctate, rugose, or combinations of both, but without an abrupt sculpture transition in lateral view; anteromedian clypeal margin subtriangular to blunt and broadly rounded: 4
4a. Head relatively longer (CI 72-76); face sculpture a combination of puncta and short transverse rugae; mandible relatively narrow with large, acute basal tooth; anteromedian clypeal margin not grossly protruding and broadly rounded: JTL-002
4b. Head relatively shorter (CI 82-87); face sculpture irregularly rugose or with large, confluent puncta; mandible relatively broad, with basal tooth angulate or blunt; anteromedian clypeal margin grossly protruding or not: 5
5a. Face sculpture irregularly rugose; basal tooth of mandible large and blunt; anteromedian clypeal margin grossly protruding and broadly rounded: paeminosa
5b. Face sculpture with large, confluent puncta underlain with granular microsculpture; basal tooth of mandible relatively small and angulate; anteromedian clypeal margin rounded but not grossly protruding: JTL-003
Key characters and variation
Mandible dentition: two strong apical teeth and one strong basal tooth are common to all. JTL-001 has highly variable dentition between the basal and subapical teeth: some have one additional strong tooth; some have two strong teeth; some have two or three small denticles; some have the basal tooth bicuspid (as though one of the denticles has moved down onto the basal tooth). pergandei is reported to have denticles between the basal and apical teeth. The remaining species have no trace of teeth or denticles between the basal and apical teeth. JTL-003 has the basal tooth somewhat enlarged and blunt relative to JTL-002, oculata, and paeminosa. paeminosa tends to have the basal tooth shifted apically relative to JTL-002 and oculata, and the tooth is shorter and relatively less acute. In JTL-003 and paeminosa the mandibles are relatively paddle-shaped, broadened medially; in JTL-002 and oculata the mandibles are sublinear.
Face sculpture: JTL-001 has transversely arcuate rugae. oculata is relatively uniformly punctate in face view (in lateral view there is a distinct transition to sublinear, transversely arcuate rugae on the ventral surface of the head). JTL-002 has puncta and short transverse rugae. JTL-003 has relatively large and confluent puncta, underlain by a granular microsculpture. paeminosa has coarse irregular rugae. pergandei is stated to be densely punctate (Wheeler 1935).
Clypeal margin: pergandei has a spine on the anterior clypeal margin. JTL-001 and oculata have a relatively acute clypeal margin. (A collection of JTL-001 from Osa Peninsula has clypeus relatively blunt, more like paeminosa.) JTL-002 and JTL-003 are transitional to paeminosa , which has a blunt anterior margin.
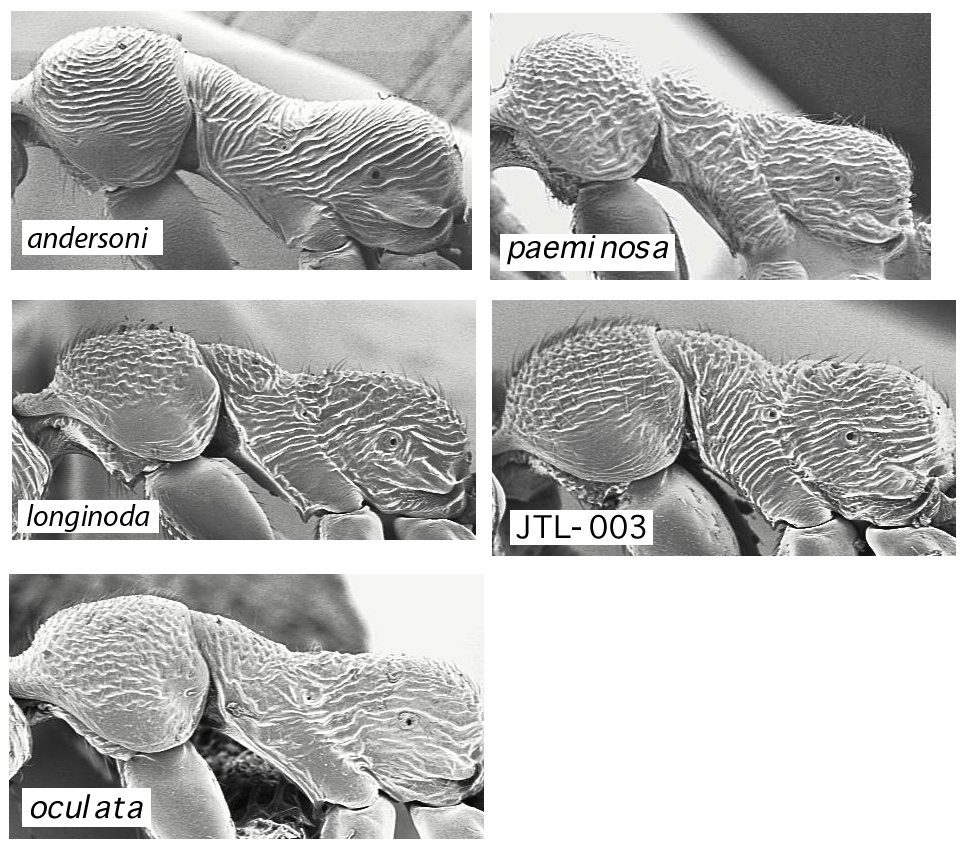
Mesosoma sculpture: JTL-001 is very distinctive, with widely-spaced sublinear striae. paeminosa is coarsely vermiculate rugose throughout, including the full extent of the lateral pronotum. JTL-002, JTL-003, and oculata are intermediate and variable, with various combinations of vermiculate and sublinear rugae, and the ventrolateral region of the pronotum is often but not always smooth and shiny. pergandei is stated to be densely punctate (Wheeler 1935).
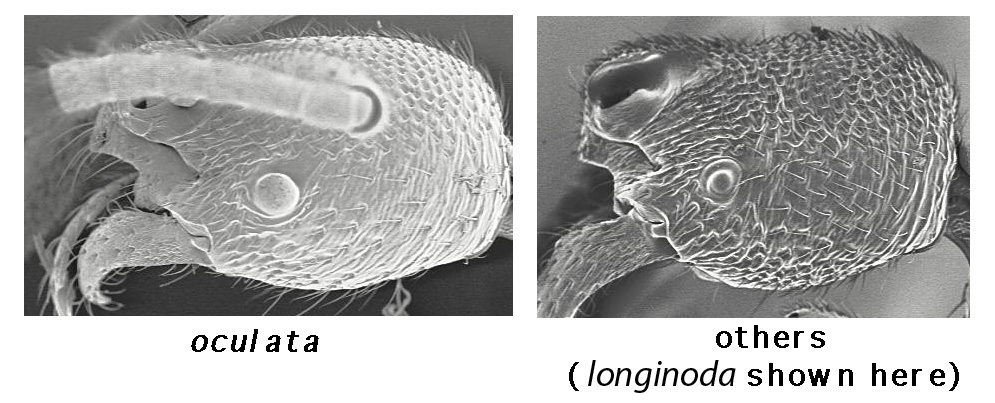
katepisternal tooth: (lateral view of mesosoma, propodeal spiracle at upper right).

Eye size:
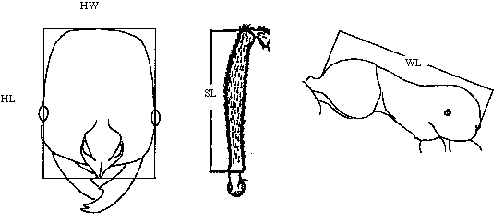
Measurements: HW = head width, measured across widest portion of head above eyes, and not including eyes; HL = head length, midline length from anterior border of clypeus to posterior border of vertex; SL = scape length; WL = Weber's length. All measurements are mm. CI = 100*HW/HL.
| Species | HL | HW | SL | WL | CI | Barcode |
|---|---|---|---|---|---|---|
| JTL-001 | 0.97 | 0.77 | 0.96 | 1.52 | 79 | INBIOCRI001279772 |
| JTL-001 | 0.97 | 0.77 | 1.00 | 1.54 | 79 | INBIOCRI001279770 |
| JTL-001 | 0.98 | 0.77 | 0.98 | 1.53 | 78 | INBIOCRI001279777 |
| JTL-001 | 1.02 | 0.80 | 0.99 | 1.56 | 78 | INBIOCRI001279776 |
| JTL-001 | 0.88 | 0.72 | 0.87 | 1.40 | 81 | INBIOCRI001279774 |
| JTL-001 | 0.97 | 0.77 | 0.97 | 1.52 | 79 | INBIOCRI001279773 |
| JTL-001 | 0.99 | 0.80 | 0.92 | 1.54 | 80 | INBIOCRI001279800 |
| JTL-001 | 0.94 | 0.72 | 0.98 | 1.51 | 77 | INBIOCRI001279798 |
| JTL-002 | 0.91 | 0.68 | 0.77 | 1.29 | 75 | INBIOCRI001279787 |
| JTL-002 | 0.85 | 0.61 | 0.74 | 1.25 | 72 | INBIOCRI001279786 |
| JTL-002 | 0.90 | 0.67 | 0.79 | 1.32 | 75 | INBIOCRI001279785 |
| JTL-002 | 0.91 | 0.69 | 0.81 | 1.32 | 76 | INBIOCRI001279783 |
| JTL-002 | 0.73 | 0.55 | 0.56 | 1.04 | 76 | INBIOCRI001279781 |
| JTL-003 | 0.86 | 0.75 | 0.76 | 1.30 | 87 | INBIOCRI001279779 |
| JTL-003 | 0.85 | 0.73 | 0.74 | 1.25 | 86 | INBIOCRI001279795 |
| JTL-003 | 0.84 | 0.72 | 0.70 | 1.23 | 86 | INBIOCRI001279802 |
| paeminosa | 0.90 | 0.74 | 0.79 | 1.37 | 82 | INBIOCRI001279794 |
| paeminosa | 0.88 | 0.74 | 0.78 | 1.36 | 84 | INBIOCRI001279792 |
| oculata | 0.89 | 0.62 | 0.80 | 1.32 | 69 | INBIOCRI001279791 |
| oculata | 1.01 | 0.73 | 0.95 | 1.56 | 72 | INBIOCRI001279790 |
| oculata | 0.94 | 0.64 | 0.82 | 1.36 | 68 | INBIOCRI001279801 |