The most conspicuous ant-plant association in the wet neotropics is that of Azteca ants and Cecropia trees. In most low-elevation, wet regions of the neotropics, Cecropia trees are ubiquitous and important invaders of man-made clearings (Uhl et al. 1981; Brokaw 1987). Their open, candelabra-shaped crowns often appear as the sole emergents over dense vine tangles in abandoned fields. The hollow internodes of these trees are almost always occupied by biting ants in the genus Azteca, each tree typically containing a single colony (Bequaert 1922; Wheeler 1942; Benson 1985).
The genus Cecropia is a highly distinctive lineage of dioecious trees, placed variously in the Cecropiaceae, Moraceae, or Urticaceae (Burger 1977, Berg 1978, Judd et al. 1994). On cladistic grounds, a broadly defined Urticaceae contains Cecropiaceae and Moraceae (Judd et al. 1994). It is native to the Neotropics, and occurs as an introduced exotic elsewhere (Ake Assi 1980; Putz and Holbrook 1988).
The basic unit of stem elongation is the internode, a cylinder bearing a single, long-petioled, palmately-lobed, peltate leaf. The internode center is filled initially with a soft pith. The pith is surrounded by a thin layer of extremely hard sclerenchyma, which is continuous across the two ends of the internode, and thus forms durable septa between the internodes (Bailey 1922). The cambium, and hence all wood growth, is external to this capsule of horny tissue.
In most of the more than 100 Cecropia species there is a suite of characters which relate to ant association (Mueller 1876, 1880-1881, Eidmann 1945). First, the internode pith splits and retracts during internode development, leaving a hollow internode with a thin layer of spongy pith on the walls. The hollow internodes form the domatia which ants inhabit. Second, each internode has a preformed thin spot, the prostoma (plural prostomata), which lacks latex ducts (Bailey 1922). The prostoma is often a well-defined oval depression in the wall, and is typically excavated and used as an entrance hole by both founding ant queens and workers of established colonies. Third, the base of each petiole bears a trichilium (plural trichilia), a pad of densely packed trichomes, from which sprout 1-2mm long glycogen-containing beads (Rickson 1971) called muellerian bodies. In Costa Rica, only the terminal two or three trichilia on leafy branches of mature trees produce muellerian bodies; production abruptly ceases on more proximal leaves. Most ant inhabitants of Cecropia harvest muellerian bodies as their primary food source.
The dominant ants associated with Cecropia are several species of Azteca (Longino 1989b, 1991b and included references). The genus Azteca is neotropical, with many species, all of which are arboreal (Emery 1893, 1896, 1912, Forel 1929, Carroll 1983). They exhibit a wide variety of arboreal nesting habits, including being obligate inhabitants of a variety of myrmecophytes. Those inhabiting Cecropia represent at least two and perhaps as many as four independent evolutionary colonizations of Cecropia (Longino 1991b, Ayala et al. 1996). Other obligate inhabitants of Cecropia include several species of Pachycondyla (Ponerinae). Although never as abundant as the dominant Azteca specialists, they are widespread in the Neotropics, and almost certainly represent multiple independent colonizations of Cecropia from within the genus Pachycondyla (pers. obs.).
Following nuptial flights, newly mated queens search out Cecropia saplings in which to initiate colonies. They chew a hole in the prostoma and enter, and plug the hole with scrapings from the internode wall. In a short time it grows in with callous tissue, sealing the queen inside. The queen founds claustrally, rearing a first brood of workers without leaving the internode.
Prior to the establishment of a colony large enough to occupy all internodes, Cecropia saplings are stacks of semi-autonomous nest sites for ants. Saplings frequently contain numerous incipient colonies in separate internodes (Longino 1989a, Davidson et al. 1991, Choe and Perlman 1997). Incipient or mature colonies of a variety of arboreal ant species may be found inhabiting lower internodes. The species list is large and includes the genera Gnamptogenys, Heteroponera, Pachycondyla, Pseudomyrmex, Crematogaster, Solenopsis, Pheidole, Wasmannia, Zacryptocerus, Procryptocerus, Camponotus, and Myrmelachista. The uppermost internodes typically contain live queens of the local species of Azteca that are obligate inhabitants of Cecropia. Internodes below those housing live queens routinely contain dead Azteca queens, evidence of repeated colony failures. These failures occur inside sealed internodes, and thus are not due to interactions with other colonies. Queens are sometimes killed by hymenopteran parasitoids (Conoaxima: Eurytomidae, see Wheeler 1942), and these may be locally abundant.
In some areas, pleometrosis (multiple-queen founding) is common, and anywhere from one to 20 or more queens, often of more than one Azteca species, may be found in the same internode (Choe and Perlman 1997).
At some point during sapling growth colonies begin to survive, producing workers that chew back out through the prostoma and begin gathering muellerian bodies from trichilia. Reduction of number of colonies occurs, such that one Azteca colony comes to dominate the sapling (Choe and Perlman 1997).
Workers restrict their movement to the host tree itself; they do not forage off the host tree. Colonies appear to derive all sustenance from the muellerian bodies, with perhaps some additional nutrition derived from scattered coccoid Homoptera on the interior stem walls.
There is debate regarding whether or not the relationship between Azteca and Cecropia is a mutualism. Workers have been observed trimming vines from the trees (Janzen 1969). One experiment demonstrated mutualism (Schupp 1986). Vasconcelos and Casimiro (1997) report that Azteca presence inhibits herbivory by Atta. Other reports question the benefit of ants for plants (Wheeler 1942, Andrade and Carauta 1982). The question should perhaps be rephrased to when and where is it a mutualism, since the species of ant occupants vary greatly in their behavior toward intruders and their use of the plant (Longino 1991a, b; species accounts below). Thus, the status of mutualism probably depends on which species of ant occupies the plant.
If not appreciated, these interspecific differences in ant behavior can also confound studies of demography (e.g. Ferguson et al. 1995). Given the strong behavioral differences, I hypothesize that the composition of the local Azteca community at the time of sapling establishment will have a profound influence on the demography of a Cecropia cohort. Demographic studies of this important component of Neotropical successional systems (e.g. Alvarez-Buylla and Martinez-Ramos 1992, Clark and Clark 1993) should also consider the role of the ants.
(most of this section is taken directly from Longino 1991b.)
Local regions support two to five Cecropia-inhabiting Azteca species. Competition appears to be intense, with multiple queens and multiple species initiating colonies in individual saplings and vying for dominance (Longino 1989a, Davidson et al. 1989, 1991, Yu and Davidson 1997). What mechanisms might lead to coexistence of species in the face of competition for the same limited resource? The answer is probably some form of niche partitioning based on preference for particular habitats and/or Cecropia species.
Azteca species are not randomly distributed with respect to habitat or Cecropia species. In at least three regions, Costa Rica, Venezuela, and Peru (Davidson, et al., 1991; Davidson and Fisher 1991, Yu and Davidson 1997), there are three-way correlations between habitat, Azteca species, and Cecropia species. Davidson and Fisher discussed possible adaptive explanations for the correlations, relying on plant defense theory. Here I discuss possible proximate mechanisms. The following causes could generate the observed patterns: 1) habitat characteristics could determine Azteca distribution, 2) habitat characteristics could determine Cecropia distribution, 3) Azteca species could influence Cecropia distribution, and 4) Cecropia species could influence Azteca distribution.
1) Habitat effect on Azteca
Habitat features include temperature, rainfall, seasonality, and disturbance regime, all of which probably influence distributions of Azteca and Cecropia. Cold, wet conditions may determine the upper elevational limit for many Azteca species. The diversity and abundance of the genus as a whole drops quickly with increasing elevation, but certain Cecropia ants are exceptional in attaining elevations of 2000m. In elevational transects, Cecropia ants always far exceed all other members of the genus. Differential tolerance of cold, wet conditions among obligate Cecropia ants could result in elevational habitat partioning based on climate alone. Examples where high elevation tolerance may promote coexistence are A. salti vs. A. constructor in Colombia, A. aragua or A. merida vs. A. lattke vs. A. constructor in Venezuela, and A. petalomyrmex vs. A. australis in Bolivia (Longino 1991b).
The long seasonal drought of tropical dry forest may have adverse affects on Cecropia ants by decreasing host-tree muellerian body production, and by decreasing internode humidity to damaging levels. Azteca coeruleipennis may be a tropical dry forest specialist, with adaptations for tolerating prolonged drought (see under A. coeruleipennis below).
Disturbance regime is probably a major factor affecting Azteca - Cecropia communities. Frequent disturbance may favor an Azteca species that reproduces early at the expense of tree defense, while infrequent disturbance may favor species with greater investment in tree defense and consequent delayed reproduction. Widely-spaced disturbances may favor the production of many small queens for long-distance dispersal. Closely-spaced disturbances may favor the production of fewer, larger queens for greater competitive ability.
I think a major pattern in Cecropia-ant communities is caused by disturbance regime. Throughout the Neotropics, A. alfari dominates Cecropia trees on frequently chopped roadsides and agricultural land, while other Azteca species dominate Cecropia trees in primary forest or very old second growth forest. Three characteristics of A. alfari favor its dominance in areas of frequent and large disturbance: early reproduction in small trees, small queen size (the smallest of the Cecropia ants), and weak defense of host tree. In contrast, most of the other species (where known) reproduce only in the boles of large trees, have larger queens, and are strong defenders of their host trees.
Azteca ovaticeps, a close relative of A. alfari, is enigmatic. It shares behavioral features and size with A. alfari, but tends to occur in areas of frequent disturbance which are embedded in primary forest: river margins in Brazil and Peru, new roadcuts or new clearings in otherwise forested areas of Costa Rica and Venezuela (Longino 1989b). Azteca ovaticeps is also more geographically variable than A. alfari. The following historical scenario would explain the current distribution and character of A. alfari and A. ovaticeps. The A. alfari group initially may have been a set of allopatric populations, each confined to small areas of high natural disturbance. One of these populations may have been particularly well-suited to a very high disturbance regime, expanding in concert with human habitat alteration in the Neotropics, and, through secondary contact, becoming sympatric with the other populations. This expanding population, now called A. alfari, would be very uniform over a large range. The remaining populations, now collectively called A. ovaticeps, would exhibit the ancestral pattern of allopatry and geographic variation.
2) Habitat effect on Cecropia
The same habitat factors which potentially influence ant distribution are likely to influence Cecropia distribution through their actions on fruit dispersal, germination, and survivorship. Cecropia species often show pronounced elevational zonation, perhaps due to climatic effects. Cecropia are probably affected by disturbance regime, some species being clearly "weedy," invading large man-made disturbances, others being "forest" species, occurring more often in gaps in primary forest.
Given that the same habitat features might influence the distribution (and segregation) of Azteca species and Cecropia species, the observed ecological "preference" (meaning correlation) of certain Azteca species for certain Cecropia species could result from independent but similar responses to these habitat features. Alternatively, biotic interactions between trees and ants might cause the species sorting and association.
3) Azteca effect on Cecropia
An ecological correlation between Azteca species and Cecropia species could result from ant effects on their host trees, in which interspecific variation in ant defensive behavior has demographic consequences for the Cecropia population. At La Selva Biological Station in Costa Rica, a strong pattern of host association is observed. Populations of small C. obtusifolia trees in young second growth vegetation, at the edge of the station property, contain the four Azteca species A. xanthochroa, A. constructor, A. alfari, and A. ovaticeps, each of them common. In contrast, large C. obtusifolia trees in old second growth forest contain only A. xanthochroa and A. constructor. This pattern may result from differential mortality of trees during succession, the relatively undefended trees inhabited by A. alfari and A. ovaticeps succumbing to herbivory and vine encroachment.
4) Cecropia effect on Azteca
Finally, correlations between tree species and ant species could result from tree effects on ants. Chemical or anatomical features of Cecropia trees could influence Azteca distribution, independent of habitat effects. Tree effects on Azteca could manifest themselves as host-specific searching by founding queens, or differential mortality of Azteca species following colonization. If either of these processes occur, they are not absolute. Founding queens are generalists with respect to species of Cecropia sapling, and will even colonize saplings of non-myrmecophytic species. Mature colonies of a single species of Azteca may be found inhabiting several Cecropia species in a region. However, tree species could still have differential effects on Azteca in a non-absolute way. Founding queens may exhibit a preference for certain tree species that is a function of motivation, such that failure to find a suitable host tree within a particular time results in addition of species to their search profile. Even if queens are absolutely generalized in their search for Cecropia saplings, fitness effects could produce patterns. Two Azteca species might have reversed competitive relationships in two Cecropia species. Each Azteca species would dominate in areas dominated by its preferred tree species, but abutting distributions or mixed populations of the two tree species would be indeterminate with respect to species of ant occupant. Elevational zonation of Azteca species could result from such a process.
At La Selva Biological Station, another feature of the pattern of host association may be explained by interspecific variation in Cecropia characteristics. In primary forest and old second growth, large C. obtusifolia trees are dominated by A. xanthochroa or A. constructor, while the intermingled population of large C. insignis trees is dominated by A. ovaticeps. Cecropia insignis appears to rely less on ant protection than C. obtusifolia. Relative to C. obtusifolia, its trichilia are more irregular and they appear later on saplings. In contrast to C. obtusifolia, young trees occupied by A. alfari and A. ovaticeps do not appear more herbivore damaged or vine-covered than those occupied by A. xanthochroa or A. constructor. Azteca ovaticeps may be favored in C. insignis because 1) C. insignis does not require strong ant defense to survive, and 2) A. ovaticeps is somehow competitively superior at colonizing C. insignis. Cecropia obtusifolia may be dominated by A. xanthochroa and A. constructor because it requires aggressive ant defense to survive, regardless of competitive relationships among ant species at colonization.
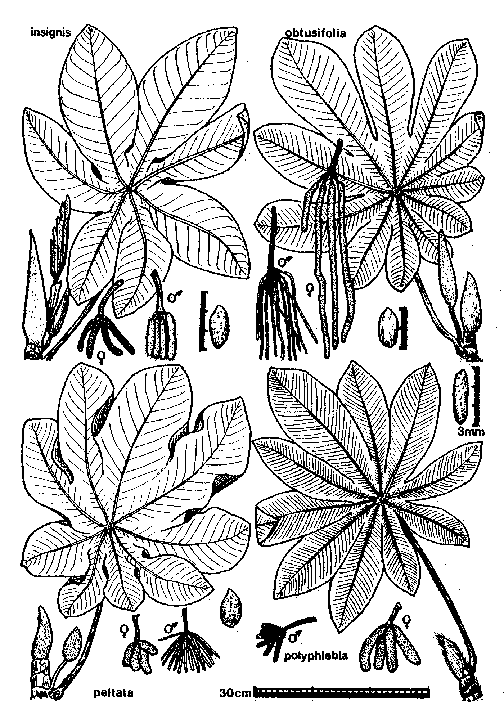
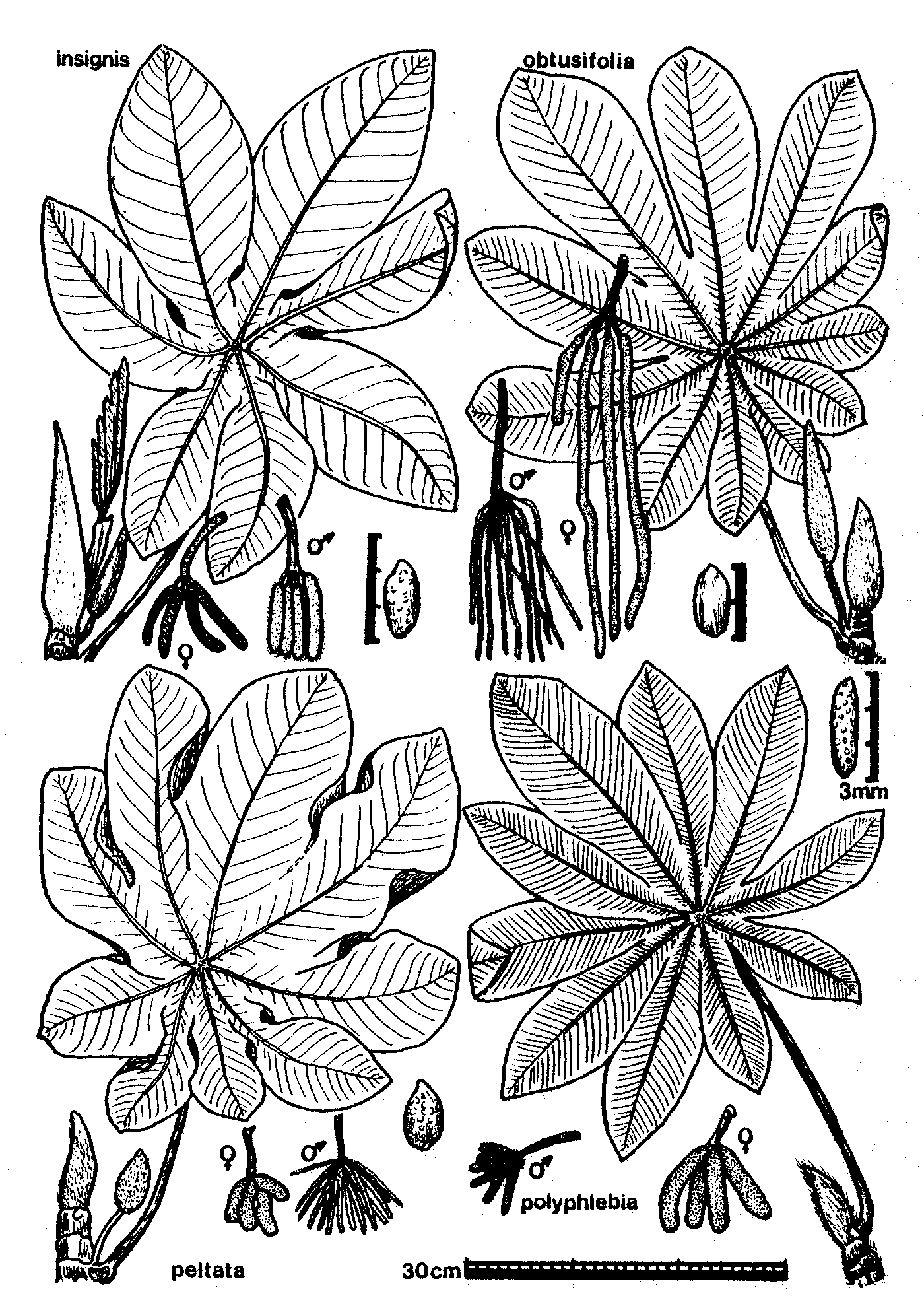
Line drawings of Costa Rican Cecropia species (from Burger 1977): 63k gif file or 189k gif file.
The Cecropia species known from mainland Costa Rica are (following Burger 1977):
Leaf surface scabrous (rough) above; usually with 9-11 lobes; longest vein of leaf with 14 to 20 pairs of major secondary veins; inflorescence spikes less than 10cm long; male spikes much thinner (2-4mm) than female spikes (5-10mm). Cecropia peltata is the species most tolerant of dry conditions in Costa Rica. It is common throughout the Pacific lowlands. Near Monteverde, C. peltata is the most common species along the road from the Pan American highway, dropping out where deciduous vegetation changes to evergreen vegetation (about 1000m elevation).
Leaf surface scabrous above; usually with 10-13 lobes; longest vein of leaf with about 30 pairs of major secondary veins; inflorescence spikes 8-22cm long in males, 18-50cm long in females; male spikes much thinner (about 1mm) than female spikes (3-5mm). Cecropia obtusifolia is common in disturbed areas and along rivers in lowland wet habitats throughout Costa Rica. Near Monteverde, C. obtusifolia is common in a narrow band on the Pacific slope, filling the lower portion of the evergreen zone (1000-1400m elevation). Below the evergreen zone C. obtusifolia is restricted to stream margins, and becomes increasingly rare at lower elevations. On the Atlantic slope east of Monteverde there is a broad gap in the distribution, with no C. obtusifolia known from at least as far down as 800m in the Penas Blancas valley. In Monteverde, C. obtusifolia intergrades with C. angustifolia (see below).
Leaf surface smooth above; leaves usually with 8-10 lobes; longest vein of leaf with 18 to 30 pairs of major secondary veins; inflorescence spikes 6-12cm long and about 1cm thick in both sexes; spikes suberect (as opposed to pendant in other species). Cecropia insignis occurs in lowland wet sites throughout Costa Rica. It is the dominant and sole Cecropia species in the very wet mid-elevation Atlantic slope forests. At lower elevations it appears restricted to older second growth forests, and is replaced by C. obtusifolia in highly disturbed areas. The trees persist to become large, long-term components of old growth forests. Near Monteverde, C. insignis begins to appear at about 1100m on the Atlantic slope, and quickly increases in abundance at lower elevations. It is essentially the only Cecropia species along the Penas Blancas river from 900m to below 800m elevation (an isolated C. angustifolia sapling was observed once near Eladio's).
Leaf surface scabrous above; usually with 10-11 lobes; longest vein of leaf with 30 to 40 pairs of major secondary veins; inflorescence spikes less than 6cm long; male spikes slightly thinner (5-8mm) than female spikes (7-16mm). Cecropia angustifolia occurs in cloud forest habitats throughout Costa Rica. Near Monteverde it is the dominant Cecropia in the cloud forest itself, being common from about 1400m on the Pacific side to 1100m on the Atlantic side. On the Pacific side it intergrades with C. obtusifolia (see below).
Cecropia peltata, C. obtusifolia, and C. insignis are conspicuously myrmecophytic. Cecropia angustifolia is non-myrmecophytic, with what appear to be vestiges of myrmecophytic characters (as though derived from a myrmecophytic ancestor; Janzen 1973). The trichilia are poorly defined and covered with long hairs. Mullerian bodies are produced, but at a very low rate. The internodes are filled with corky brown pith. Mature trees are never inhabited by Azteca (although Myrmelachista, Procryptocerus, and Camponotus may be found nesting opportunistically in the stems). Saplings frequently contain founding Azteca queens (presumably having flown up from lower elevations), even though they never survive to establish colonies. Azteca queens apparently do not perceive C. angustifolia as an unsuitable host.
Cecropia obtusifolia and C. angustifolia appear to intergrade in Monteverde. "Typical" C. obtusifolia occurs in the upper San Luis valley and the steep slopes just below the Monteverde community. "Typical" C. angustifolia occurs at the upper edge of the community and in the Reserve. In between is a continuous gradation in leaf shape, shape of male and female reproductive structures, and development of myrmecophytic characters. Obligate plant ants drop out in this transition zone, at the level of the Monteverde meeting house. This same gradation between C. obtusifolia and C. angustifolia occurs in the Las Alturas area in the Talamanca range in southern Costa Rica (pers. obs.).
Earlier reports refer to Costa Rican populations of this species as C. polyphlebia. Berg and Franco Rosselli (2005) synonymized C. polyphlebia under C. angustifolia.
Scapes and tibiae with abundant conspicuous setae; queen orange or mottled orange brown; workers generally brown, largest workers approaching queen in color, with orange laterally on face; head width of largest workers approaching 1.4mm.
Azteca xanthochroa is common in wet zones throughout Costa Rica. It is usually abundant in or near old second growth vegetation, where Cecropia trees are allowed to grow to large size. In the Atlantic lowlands, it is common in mature C. obtusifolia trees. At higher elevations on the Atlantic slope, such as the Penas Blancas Valley, it is common in mature C. insignis trees. It is the most common species in the band of C. obtusifolia that is found in and just below Monteverde on the Pacific slope. Descending westward from Monteverde, its abundance declines abruptly, corresponding to the abrupt transition from dominance of C. obtusifolia to dominance of C. peltata (Longino 1989a), and from wet conditions to dry conditions. Although its distribution correlates with C. obtusifolia, it is not due to a strict host specificity: colonies can be found in C. peltata at the transition from wet to dry habitats (Longino 1989a).
Colonies have a single cylindrical carton nest in the bole of the tree, and the colony queen, all larvae, and any alate sexuals are concentrated in this single nest. All the septa between the nest and the branch tips are perforated, so that there are continuous internal passages between nest and branch tips. Unlike all other Cecropia ants, A. xanthochroa workers maintain a vertical fissure near the base of the tree, well below the carton nest, and they do not maintain fissures near the nest itself. The fissure is usually less than two or three meters from the ground, even in large trees. When the tree is disturbed, large workers emerge from the fissure, and many workers pour out of holes at the branch tips. The workers are aggressive and rapidly attack intruders on the foliage (Longino 1991a).
Scapes and tibiae with abundant, conspicuous setae; queen black with dense brush of setae on head and mesosomal dorsum; workers brown, difficult to distinguish from small workers of A. xanthochroa; head width of largest workers approaching 1.4mm.
Azteca constructor occurs throughout Costa Rica, wherever myrmecophytic Cecropia occur. It is similar to A. xanthochroa in favoring old second growth vegetation where Cecropia trees attain large size. It has a broader tolerance of moisture conditions, being found commonly in both wet and dry forest. It can be found regularly in all three of the myrmecophytic Cecropia species in Costa Rica. It is common in the Monteverde area, on both slopes and in the community area.
Mature colonies occupy a single carton nest in the bole of the tree, where brood and reproduction are concentrated. The nest is spindle-shaped and often causes a deformation of the trunk. Many entrance holes and fissures are maintained near the carton nest itself, and internal passages are maintained to all the branch tips. Workers of this species are extremely aggressive, and respond to any disturbance by pouring out of large fissures near the carton nest and blackening the trunk surface (Longino 1991a).
Scapes and tibiae with abundant but inconspicuous setae; queen shiny black, without the conspicuous brush of setae of A. constructor; workers uniformly light yellow orange; head width of largest workers less than 1.05mm; rear part of mesosoma flattened and shelf-like.
Azteca coeruleipennis is common in dry forest areas of Costa Rica and exhibits abrupt declines at habitat boundaries (Longino 1989a). It occurs throughout the Guanacaste lowlands south to the central valley and adjacent Pacific coastal areas. It usually inhabits C. peltata but may occur in C. obtusifolia and C. insignis at habitat boundaries where these species become more abundant. Near Monteverde, A. coeruleipennis is common along the road from the PanAmerican highway and in the San Luis valley, abruptly dropping out at about 900m elevation.
Mature colonies have a dispersed colony structure. There is no central carton nest. Brood and any alate sexuals are in branch tips. Older parts of the tree are gradually abandoned, and internal passages are not maintained among occupied branches. When a tree is molested bright yellow workers of rather uniform small size issue forth and descend the trunk, where they aggressively attack the intruder.
Azteca coeruleipennis may be a tropical dry forest specialist, with adaptations for tolerating prolonged drought. During the dry season, leafy branch tips are perforated by one or two active entrances, often 10 or more internodes back from the terminal sheathing stipule. In contrast, other Azteca species typically maintain five or more active entrances in the terminal 10 internodes (pers. obs.). Occupied branch sections which contain alate queens have active entrances which are only large enough for workers. Larger tunnels are excavated in the internode walls, but these end blindly within a millimeter of surface, as though the final perforation is suspended until wet season and/or the time of flight of new queens. Reduction of the number and size of entrances may be an adaptation for reducing water loss (Longino 1991a).
Scapes and tibiae devoid of setae; queen black to variously red brown and smaller than any of the above species.
Members of the A. alfari complex occur throughout Costa Rica, and will inhabit any of the myrmecophytic Cecropia species. They generally prefer open and highly disturbed habitats, and may reproduce in relatively small and short-lived trees. In the Monteverde area, they parallel the distribution of A. coeruleipennis. They are extremely rare in the Penas Blancas valley and in the community area, and they increase in abundance as one descends the Pacific slope. They are very common in Cecropia trees along the lower San Luis River.
Mature colonies of the A. alfari complex have a dispersed structure like that of A. coeruleipennis. Unlike the above species, workers are relatively timid and will not defend a mature tree (small saplings are more aggressively defended). When a tree is disturbed, workers usually remain in the branches. Only when a branch is broken open will workers rush out to attack. Trees occupied by members of the A. alfari complex often appear somewhat decrepid, with abundant leaf damage and frequently burdened with vines. The foliage of these trees often hosts a diverse insect fauna, including foraging ants of other species.
In many parts of the Neotropics there are two locally sympatric forms in the A. alfari complex, one of which is less setose than the other. Longino (1989b) treated them as two species, with the less setose species being A. alfari and the more setose species being A. ovaticeps. The difference in pilosity can be quantified on queens by counting the number of setae on the second tergite (top half of the segment) of the gaster (abdomen beyond petiole), not including the row of setae on the posterior border. Azteca alfari has 0-3 setae, and A. ovaticeps has more than 10. The distinction is often clear when members of the A. alfari complex are locally abundant. For example, in the Atlantic lowlands of Costa Rica, A. ovaticeps queens have a dense brush of setae on the abdomen, and are conspicuously distinct from A. alfari in the same area. However, in some areas the distinction is not as clear. Monteverde is one of those areas. Some colonies are identical to standard A. alfari s.s. from elsewhere in Costa Rica. Other colonies show a range of seta abundances, but are not as densely setose as the Atlantic lowland A. ovaticeps.
Pachycondyla insignis (Brown manuscript name) is an undescribed species known only from Cecropia saplings. This large ponerine is similar to the widespread and generalized P. villosa, but has transverse rugae on the clypeus. Pachycondyla villosa has the clypeus smooth or with longitudinal rugae. Pachycondyla insignis makes specialized use of Cecropia, chewing large circular entrance holes through the prostomata, and foraging on mullerian bodies (pers. obs.). They have never been collected away from a Cecropia sapling. The species is known from various sites throughout the Atlantic lowlands of Costa Rica. Occurrences near Monteverde include several collections from C. insignis saplings in the Penas Blancas valley, and one collection from a C. obtusifolia sapling between Santa Elena and Monteverde.
In addition to these obligate Cecropia ants, the weak plant ant Azteca beltii has been observed once in a Cecropia tree below Santa Elena.
Ake Assi, L. 1980. Cecropia peltata Linne (Moracees): ses origines, introduction et expansion dans l'est de la Cote d'Ivoire. Institut Fondamental d'Afrique Noire. Bulletin. Serie A. Sciences Naturelles 42:96-102.
Alvarez-Buylla, E. R., and M. Martinez-Ramos 1992. Demography and allometry of Cecropia obtusifolia, a neotropical pioneer tree - an evaluation of the climax-pioneer paradigm for tropical rain forests. Journal of Ecology 80:275-290.
Andrade, J. C. de, and J. P. P. Carauta. 1982. The Cecropia-Azteca association: a case of mutualism? Biotropica 14:15.
Ayala, F. J., J. K. Wetterer, J. T. Longino, and D. L. Hartl. 1996. Molecular phylogeny of Azteca ants (Hymenoptera: Formicidae) and the colonization of Cecropia trees. Molecular Phylogenetics and Evolution 5:423-428.
Bailey, I. W. 1922. Notes on neotropical ant-plants I. Cecropia angulata, sp. nov. Botanical Gazette 74:369-391.
Benson, W. W. 1985. Amazon ant-plants. in Amazonia, eds. G. T. Prance and T. E. Lovejoy, 239-266. Oxford, England: Pergamon.
Bequaert, J. 1922. Ants of the American Museum Congo expedition. IV. Ants in their diverse relations to the plant world. Bulletin of the American Museum of Natural History 45:333-583.
Berg, C. C. 1978. Cecropiaceae, a new family of Urticales. Taxon 27:39-44.
Berg, C. C., and P. Franco Rosselli. 2005. Cecropia. New York Botanical Garden Press, New York, USA.
Brokaw, N. V. L. 1987. Gap-phase regeneration of three pioneer tree species in a tropical forest. Journal of Ecology 75:9-19.
Burger, W. 1977. Flora Costaricensis, Family #52, Moraceae. Fieldiana, Botany 40:94-215.
Carroll, C. R. 1983. Azteca (hormiga Azteca, Azteca ants, Cecropia ants). Pages 691-693 in D. H. Janzen, editor. Costa Rican natural history. University of Chicago Press, Chicago, Illinois, USA.
Choe, J. C., and D. L. Perlman. 1997. Social conflict and cooperation among founding queens in ants (Hymenoptera: Formicidae). Pp. 392-406 in J. C. Choe and J. Crespi, eds. The evolution of social behavior in insects and arachnids. Cambridge University Press, Cambridge, England.
Clark, D. B., and D. A. Clark. 1993. Comparative analysis of microhabitat utilization by saplings of nine tree species in neotropical rain forest. Biotropica 25:397-407.
Davidson, D. E., Snelling, R. R., and Longino, J. T., 1989. Competition among ants for myrmecophytes and the significance of plant trichomes. Biotropica 21: 64-73.
Davidson, D. E., and B. L. Fisher. 1991. Symbiosis of ants with Cecropia as a function of light regime. Pages 289-309 in C. R. Huxley and D. F. Cutler, editors. Ant-plant interactions. Oxford University Press, Oxford, U.K.
Davidson, D. W., R. B. Foster, R. R. Snelling, and P. W. Lozada. 1991. Variable composition of some tropical ant-plant symbioses. Pages 145-162 in P. W. Price, T. M. Lewinsohn, G. W. Fernandes, and W. W. Benson, editors. Plant-Animal interactions: evolutionary ecology in tropical and temperate regions. John Wiley & Sons, Inc., New York.
Eidmann, H. 1945. Zur Kenntnis der Okologie von Azteca muelleri Em. (Hym. Formicidae), ein Beitrag zum Problem der Myrmecophyten. Zoologische Jahrbuecher, Abteilung fuer Systematik, Okologie und Geographie der Tiere 77:1-48, 3 tables.
Emery, C. 1893. Studio monografico sul genere Azteca Forel. Memorie della Royal Accademia delle Scienze dell'Istituto di Bologna (5)3:119-152.
Emery, C. 1896. Alcune forme nouve del genere Azteca For. e note biologiche. Bollettino dei Musei di Zoologia ed Anatomia comparata della R. Universita di Torino 11(230):1-7.
Emery, C. 1912. Subfam. Dolichoderinae. Genera Insectorum Fasc. 137:1-50.
Ferguson, B. G., D. H. Boucher, M. Pizzi, and C. Rivera. 1995. Recruitment and decay of a pulse of Cecropia in Nicaraguan rain forest damaged by Hurricane Joan: relation to mutualism with Azteca ants. Biotropica 27:455-460.
Forel, A. 1929. The social world of the ants (C. K. Ogden, Trans.). Albert and Charles Boni, New York.
Janzen, D. H. 1969. Allelopathy by myrmecophytes: the ant Azteca as an allelopathic agent of Cecropia. Ecology 50:147-153.
Janzen, D. H. 1973. Dissolution of mutualism between Cecropia and its Azteca ants. Biotropica 5:15-28.
Judd, W. S., R. W. Sanders, and M. J. Donoghue. 1994. Angiosperm family pairs: preliminary phylogenetic analyses. Harvard papers in Botany 5:1-51.
Longino, J. T. 1989a. Geographic variation and community structure in an ant-plant mutualism: Azteca and Cecropia in Costa Rica. Biotropica 21:126-132.
Longino, J. T. 1989b. Taxonomy of the Cecropia-inhabiting ants in the Azteca alfari species group: evidence for two broadly sympatric species. Contributions in Science (Natural History Museum of Los Angeles County) 412:1-16.
Longino, J. T. 1991a. Azteca ants in Cecropia trees: taxonomy, colony structure, and behavior. Pages 271-288 in C. Huxley and D. Cutler, editors. Ant-plant interactions. Oxford University Press, Oxford, U.K.
Longino, J. T. 1991b. Taxonomy of the Cecropia-inhabiting Azteca ants. Journal of Natural History 25:1571-1602.
Mueller, F. 1876. Ueber das Haarkissen am Blattstiel der Imbauba (Cecropia), das Gemuesebeet der Imbauba-Ameise. Jenaische Zeitschrift fur Medizin und Naturwissenschaft 10:281-286.
Mueller, F. 1880-1881. Die Imbauba und ihre Beschuetzer. Kosmos 8:109-116.
Putz, F. E., and N. M. Holbrook. 1988. Further observations on the dissolution of mutualism between Cecropia and its ants: the Malaysian case. Oikos 53:121-125.
Rickson, F. R. 1971. Glycogen plastids in Mullerian body cells of Cecropia peltata - a higher green plant. Science 173:344-347.
Schupp, E. W. 1986. Azteca protection of Cecropia: ant occupation benefits juvenile trees. Oecologia 70:379-385.
Uhl, C., K. Clark, H. Clark, and P. Murphy. 1981. Early plant succession after cutting and burning in the upper Rio Negro region of the Amazon basin. Journal of Ecology 69:631-649.
Vasconcelos, H. L., and A. B. Casimiro. 1997. Influence of Azteca alfari ants on the exploitation of Cecropia trees by a leaf-cutting ant. Biotropica 29:84-92.
Wheeler, W. M. 1942. Studies of neotropical ant-plants and their ants. Bulletin of the Museum of Comparative Zoology, Harvard 90:1-262.
Yu, D. W., and D. W. Davidson. 1997. Experimental studies of species-specificity in Cecropia-ant symbioses. Ecological monographs 67:273-294.
Longino, J. T. 1989a. Geographic variation and community structure in an ant-plant mutualism: Azteca and Cecropia in Costa Rica. Biotropica 21:126-132.
(PDF)
Longino, J. T. 1989. Taxonomy of the Cecropia-inhabiting ants in the Azteca alfari species group: evidence for two broadly sympatric species. Contributions in Science (Natural History Museum of Los Angeles County) 412:1-16.
(PDF)
Longino, J. T. 1991a. Azteca ants in Cecropia trees: taxonomy, colony structure, and behavior. Pages 271-288 in C. Huxley and D. Cutler, editors. Ant-Plant Interactions. Oxford University Press, Oxford. (PDF)
Longino, J. T. 1991b. Taxonomy of the Cecropia-inhabiting Azteca ants. Journal of Natural History 25:1571-1602. (PDF)
Longino, J. T. 1996. Taxonomic characterization of some live-stem inhabiting Azteca (Hymenoptera: Formicidae) in Costa Rica, with special reference to the ants of Cordia (Boraginaceae) and Triplaris (Polygonaceae). Journal of Hymenoptera Research 5:131-156. (PDF)
Page author:
John T. Longino, The Evergreen State College, Olympia WA 98505
USA. longinoj@evergreen.edu
Date of this version: 22 September 2005.
Previous versions of this page:
 Go back to top
Go back to top





{kind=link}
{kind=link}