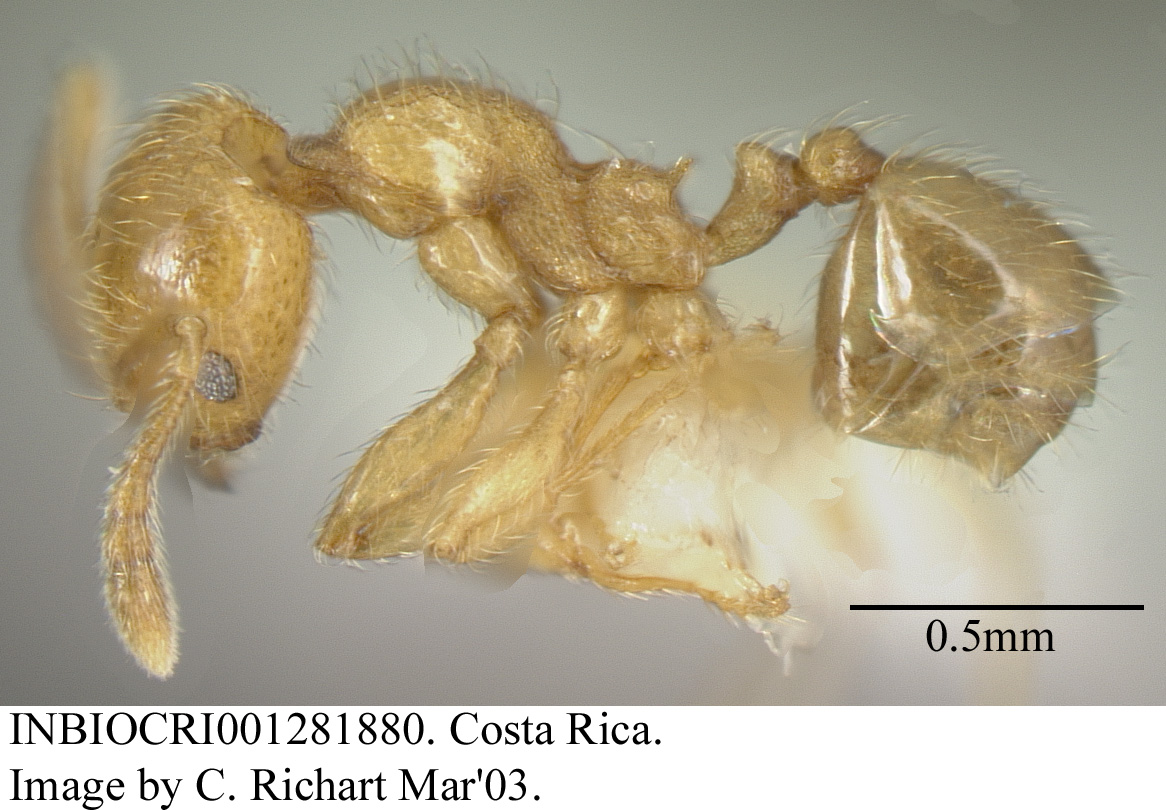


Formicidae, Hymenoptera, Insecta, Arthropoda, Animalia
|
|
|
Atlantic lowlands |
Pacific lowlands |
Identification
Minor worker (La Selva): head length 0.54mm, head width 0.47mm, scape length 0.39mm, Webers length 0.67mm (n=1). Head somewhat flattened behind; promesonotum evenly arched, mesonotal suture absent; propodeal spines small but distinct; face smooth and shining, pronotum feebly punctatorugose dorsally, with large smooth and shiny patches on sides, mesonotum and propodeum foveolate; dorsal pilosity abundant, short, suberect, flexuous; color orange.
Major worker (La Selva): head length 1.04mm, head width 0.78mm, scape length 0.42mm (n=1). Anterior margins of frontal carinae in the form of distinct teeth, elevated and projecting from face; face entirely and coarsely punctatorugose, vermiculate rugae obliquely oriented in middle, irregular on sides, and transverse on vertex lobes; hypostomal margin nearly flat, with pair of sharp teeth located about one third distance from midline to recessed teeth flanking mandibles; dorsal pilosity abundant, short, somewhat lanose; sides of head with appressed to subdecumbent short pilosity, without conspicuous erect setae.
Variation: major workers from the Pacific side of Costa Rica have relatively wider heads (head length 1.06mm, head width 0.91mm, n=1), the hypostomal margin more concave, and the hypostomal teeth much smaller (n = 4 Pacific slope collections, 3 Atlantic slope collections).
Range
Panama (type locality Bugaba), Costa Rica (Atlantic and southern Pacific lowlands).
Natural History




Pheidole bicornis is one of only two Pheidole species known to be obligate plant ants (the other being Pheidole minutula, associated with a variety of ant plants in South America). Pheidole bicornis has developed a coevolved association with a group of Piper species in the wet forests of Costa Rica and adjacent portions of Panama and Nicaragua (Fischer et al. 2002, 2003; Letourneau 1983, 1990, 1991, 1998; Rickson and Risch 1984; Risch 1982; Risch et al. 1977). The pipers always contain nests of P. bicornis, and the ant species is never found nesting elsewhere. In Costa Rica the plants are common in wet forests of both Atlantic and Pacific slopes, to about 700m elevation.
Information on the species of ant pipers in Costa Rica can be found in Burger (1971), Tebbs (1989), and Tepe et al. (2004). Species of obligate ant pipers in Costa Rica are P. calcariformis, P. cenocladum, P. fimbriulatum, P. obiquum, P. sagittifolium. Four of the species are figured below, from Burger (1971) and Tepe et al. (2004).
Judging from specimen databases at INBio and Missouri Botanical Garden, Piper cenocladum occurs throughout the country but is most abundant on the Atlantic slope. It is the common species at La Selva Biological Station. Piper fimbriulatum occurs in mid to low elevation wet forest sites in all the cordilleras, the southern Pacific lowlands, and the southern Atlantic lowlands, but is absent from the northern Atlantic lowlands. Piper obliquum occurs throughout the country. Piper sagittifolium is largely restricted to the southern Pacific slopes. Piper calcariformis occurs at mid-elevation on the Atlantic slope.
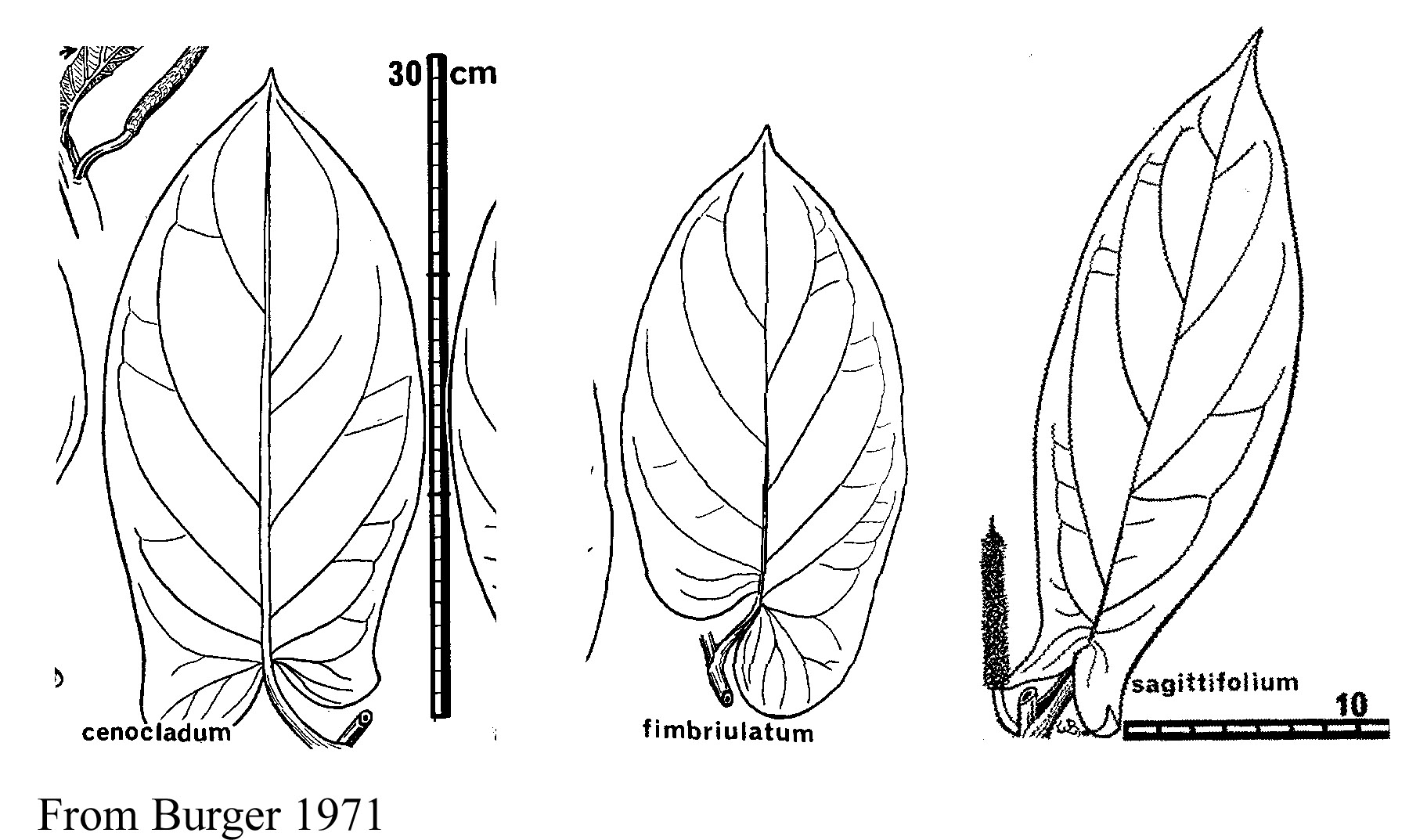
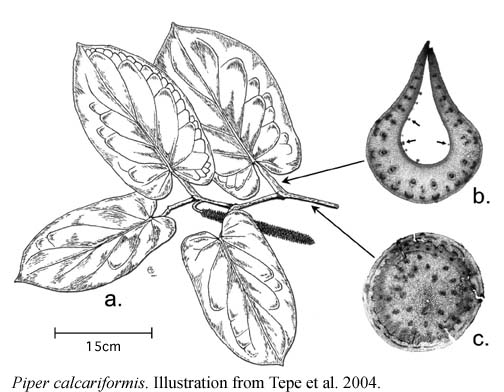
The pipers are 1-2m tall plants that grow in the shaded understory of mature rainforest. The leaf petioles have expanded margins that are "clasping" such that they form an enclosed chamber in which the ants nest. All of the ant pipers except P. calcariformis also have hollow stems or soft medullar pith that the ants excavate, with passageways from the petiolar chamber into the hollow stems. Stems of P. calcariformis are solid (Tepe et al. 2004). The colony is distributed throughout multiple petiolar chambers and hollow stems of the plant. The plants produce small white food bodies (pearl bodies) on the inner walls of the petiolar chambers and these appear to be the main food source of the ants (Fischer et al. 2002). Food body production is induced by the ants because plants grown in greenhouses without ants do not produce food bodies. Workers do not forage off their host plant, but they do forage at a low rate on the leaf surfaces.
Images of nests in Piper ant plants: click here.
Colonies are often polydomous, with multiple nests in different stems of one plant or adjacent stems of multiple plants. Fischer et al. (2003) reported a single colony in adjacent stems of two different Piper species. Colonies have dozens to hundreds of workers. A colony I observed in Corcovado National Park contained 45 major workers, 70 minor workers, 7 adult males, 1 dealate queen, and brood. Colonies appear to invest heavily in major workers, maintaining a relatively high percentage of majors in the worker force. Most colonies appear to be monogynous, but I observed one in Carara Biological Reserve (Bijagual) that was polygynous.
The Pheidole-Piper association has been extensively investigated by Letourneau. She discovered that even though the ants are relatively passive and forage at a low rate, they still provide a net benefit to the plant by removing herbivore eggs and other small herbivores (Letourneau 1983). In subsequent studies she discovered a beetle in the family Cleridae that is a parasite of this mutualism, living in the petiolar chambers and feeding on ant larvae and plant food bodies (Letourneau 1990, 1991).
Fischer et al. (2003) studied nitrogen transfer from ants to plants, and their summary follows:
1 Nutrient fluxes play a significant role in the interaction of myrmecophytic plants and their symbiotic ants. There is a clear flux from plants to ants via nectar or food bodies but nutrient fluxes from ants to plants are less obvious.
2 We report on a nitrogen flux from ants to plants in the association between Pheidole bicornis (Formicidae-Myrmicinae) and two myrmecophytic Piper species (P. fimbriulatum and P. obliquum, Piperaceae). Pulse experiments were performed by feeding ants with 15N-labelled glycine supplied in sucrose solution. Workers passed ingested label on to other ants and the brood by trophallaxis.
3 The distribution of label within the colony showed highest incorporation rates in larvae and in the working caste, while the reproductive caste received only a small amount of the 15N-labelled food.
4 Nutrient transfer from ants to plants occurred remarkably fast. Within 6 days, up to 25% of the nitrogen ingested by the ants was incorporated by the plants.
5 However, ant distribution within P. fimbriulatum plants did not correlate with the intra-plant uptake pattern of 15N, and ant-mediated nitrogen uptake by myrmecophytic P. fimbriulatum accounted for less than 1% of the plants' above-ground nitrogen demand.
Tepe et al. (2004) carried out a phylogenetic analysis of Piper section Macrostachys, which contains all the obligate ant pipers. The section also contains numerous species that Tepe et al. interpret as facultative ant plants, and a number of non-ant plants. Facultative ant plants have partially to completely developed petiolar chambers, no or sparse pearl bodies, and solid stems. Various opportunistic arboreal ants can be found nesting in them. The phylogenetic analysis revealed that obligate myrmecophytism with Pheidole bicornis evolved independently two to four times. Piper cenocladum is the most distantly related to the other four Costa Rican myrmecophyte species, and its relationship with P. bicornis clearly evolved independently of the others.
The Pheidole-Piper association is the subject of much current research and it is becoming a model system in studies of community ecology (Dyer and Palmer 2004).
Literature Cited
Burger, W. 1971. Flora Costaricensis, Family #41, Piperaceae. Fieldiana, Botany 35:5-227.
Dyer, L. A., and A. D. N. Palmer, editors. 2004. Piper: a model genus for studies of phytochemistry, ecology, and evolution. Kluwer Academic Publishing, New York.
Fischer, R. C., A. Richter, W. Wanek, and V. Mayer. 2002. Plants feed ants: food bodies of myrmecophytic Piper and their significance for the interaction with Pheidole bicornis ants. Oecologia 133:186-192.
Fischer, R. C., Wanek, W., Richter, A. and Mayer, V. 2003. Do ants feed plants? A 15N labelling study of nitrogen fluxes from ants to plants in the mutualism of Pheidole and Piper. Journal of Ecology 91:126-134.
Letourneau, D. K. 1983. Passive aggression: an alternative hypothesis for the Piper-Pheidole association. Oecologia 60:122-126.
Letourneau, D. K. 1990. Code of ant-plant mutualism broken by parasite. Science 248:215-217.
Letourneau, D. K. 1991. Parasitism of ant-plant mutualisms and the novel case of Piper. Pp. 390-396 in C. Huxley and D. Cutler, eds. Ant-Plant Interactions. Oxford University Press, Oxford.
Letourneau, D. K. 1998. Ants, stem-borers, and fungal pathogens: experimental tests of a fitness advantage in Piper ant-plants. Ecology 79:593-603.
Rickson, F. R., and S. J. Risch. 1984. Anatomical and ultrastructural aspects of the ant-food cell of Piper cenocladum C. DC. (Piperaceae). American Journal of Botany 71:1268-1274.
Risch, S. 1982. How Pheidole ants help Piper plants. Brenesia 19:545-548.
Risch, S., M. McClure, J. Vandermeer, and S. Waltz. 1977. Mutualism between three species of tropical Piper (Piperaceae) and their ant inhabitants. American Midland Naturalist 98:433-444.
Tebbs, M. C. 1989. Revision of Piper (Piperaceae) in the New World 1. Review of characters and taxonomy of Piper section Macrostachys. Bulletin of the British Museum (Natural History). Botany 19:117-158.
Tepe, E., M. Vincent, and L. Watson. 2004. Phylogenetic patterns, evolutionary trends, and the origin of ant-plant associations in Piper section Macrostachys: Burger's hypothesis revisited. Pages 156-178 in L. A. Dyer and A. D. N. Palmer, editors. Piper: a model genus for studies of phytochemistry, ecology, and evolution. Kluwer Academic Publishing, New York.
Page authors:
John T. Longino, The Evergreen State College, Olympia WA 98505 USA. longinoj@evergreen.edu
Stefan Cover, Museum of Comparative Zoology, Harvard University, Cambridge MA 02138 USA. scover@oeb.harvard.edu
Date of this version: 8 January 2005.

